单位:上海光语生物科技有限公司
微信公众号:Leadingtec
作者:俞建中(微信号:Scophy117)
俞建中博士系列藻类科普文章《光语带你认识微藻》第一辑链接:
俞建中博士系列藻类科普文章《光语带你认识微藻》第二辑链接:
光语带你认识微藻(第二辑) 第一篇 共生藻之虫黄藻
1.颗石藻的基本特征
颗石藻(Coccolithophores),又称球石藻,是一类单细胞海洋微藻,为鞭毛藻的一类(但有些能游动,有些不能游动),细胞覆盖着一层钙质外壳,又成为钙板金藻。颗石藻含叶绿素a和叶绿素c,Ehrenberg 1836年在波罗的海首先发现,而后各种不同的类型在全世界都有发现,是一类广布的海洋藻类,几乎遍布在阳光可及的所有大洋洋面。
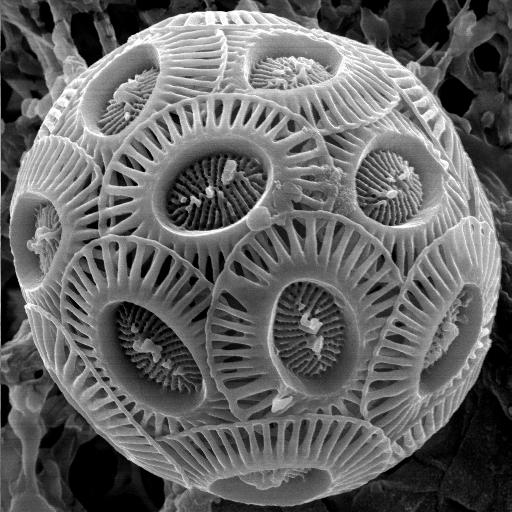
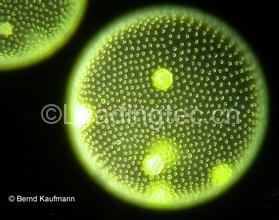
图1. 赫氏颗石藻Emiliania huxleyi ,目前分布最广泛的颗石藻种类
颗石藻的外形非常独特,有许多小碟片(鳞片),6-30片或者更多的数量,包围在细胞生物质团外面,形成球形的细胞外骨架,这种小碟片称为颗石(coccoliths),质地为碳酸钙(方解石态)、具有独特花纹和形状结构。当细胞死亡的时候,碳酸钙的外壳得以保留,大量的沉积在水底矿化后就成为白色的岩层——“白垩”。颗石藻类的个头大小不一,直径从5到100微米散布,而颗石的穿面大小从2到25微米不等。每个细胞有两个褐色的色素体,包围在细胞核外。
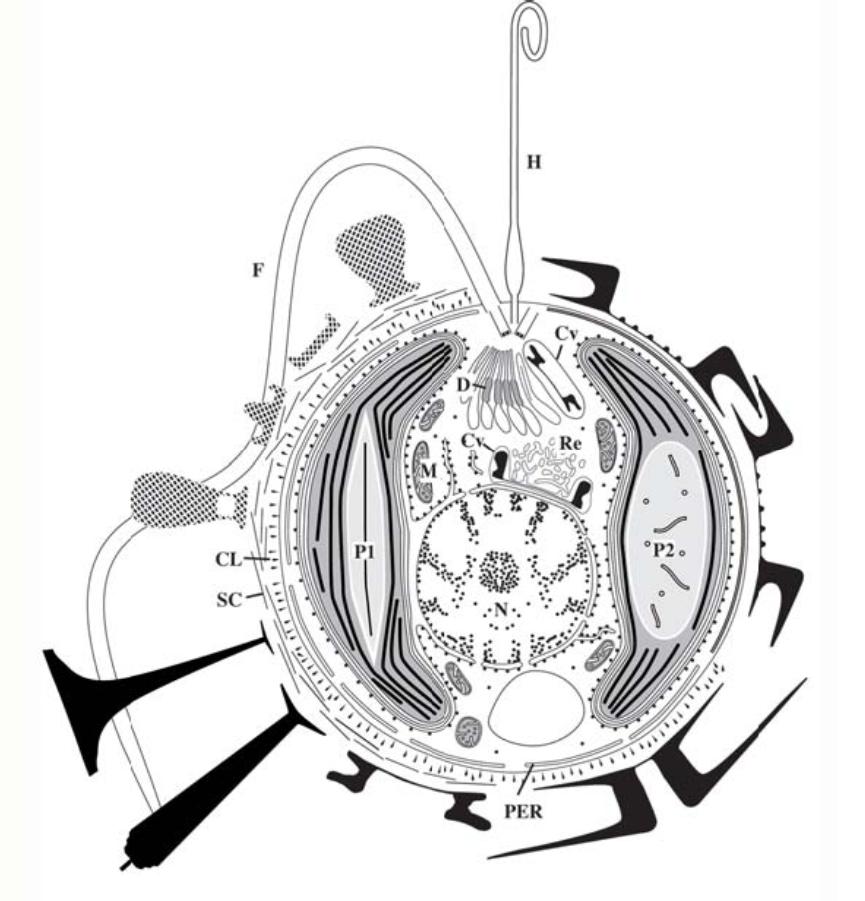
图2. 颗石藻细胞结构示意图(根据多种颗石藻特征综合而成)。P1为颗石藻中典型的细胞器“蛋白核”,而蛋白核P2在Emiliania和 Gephyrocapsa中发现。细胞外围黑色标记示意为异晶颗石,密点标记示意为同晶颗石。CL:柱状胚、Cv:形成颗石的细胞器、D:高尔基体的扩大部位;F:鞭毛;H:附着鞭毛;M:线粒体;N:细胞核;P1:类囊体贯穿的蛋白核;P2:微管贯穿的蛋白核;PER:外围内质网;Re:网状体;SC:未矿化的有机屑。(Billard & Innouye, 2004)
2.分类
(微藻的分类一直是个痛苦的事情,在此不深究,只是简单选个别专家的观点,知道是怎么回事就行)
在分类上,依据目前较新的生物分类系统,颗石藻属于定鞭藻类的一类,属普林藻纲(Prymnesiophyceae,有称定鞭藻纲),也有分出个小纲叫颗石藻(亚)纲(Coccolithophyceae),(定鞭藻一般认为由巴夫藻纲Pavlovophyceae和普林藻纲Prymnesiophyceae两个大纲组成)。
首先定鞭藻类跟其他真核生物之间的关系就搞不清,跟一个叫“SAR”的组合还接近点 [不等鞭毛生物(Stramenopiles),囊泡虫(Alveolates),有孔虫(Rhizaria),一起称为“SAR”组合]。
普林藻纲中许多种类不具有形成颗石的能力,它的特点是具有两根等长或接近等长的光滑的鞭毛,有些种类细胞体覆盖了有机或矿物质。普林藻纲中也是花样各异,上述特征并不能覆盖所有种类,下分有六个目,又有些种类没法归属到这些目中,而颗石藻就是所有种类中有“颗石”的种类的统称(有同行就碰到“颗石藻”没颗石的情况,可能这颗石就不能作为分类的依据,例如有些人多个指头的情况)。有意思的是,普林藻类形态描述中细胞体覆盖矿物质,所谓的矿物质,钙质是碳酸钙形式,但有方解石态(calcite)和文石态(aragonite)两种,除了钙质种类还有硅质的种类,当然它们不是硅藻。
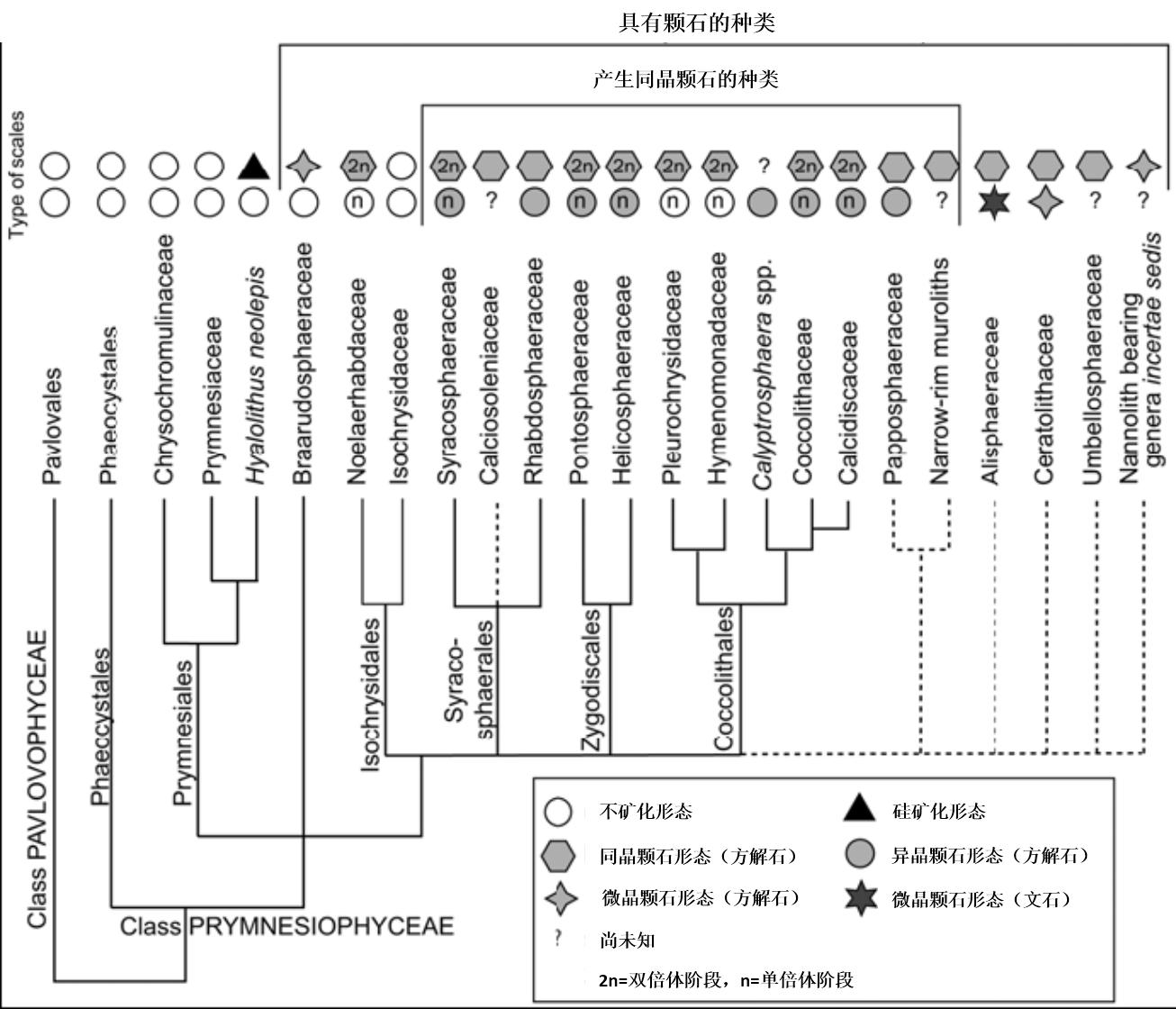
图3. 不同种类的亲缘关系就不解释了,种名也不翻译了,汉字虽小倒是值得注意,图中统计了巴夫藻纲和普林藻纲中可称为颗石藻的种类,及其他一些已发现的特征,如颗石类型等(Hagino & Young,2015)
3. 颗石藻与颗石形态
颗石藻通常根据藻体的形态和其颗石的晶体结构来进行分类。颗石通常可以分为三类,异晶颗石(Heterococcoliths, HET)、同晶颗石(Holococcoliths, HOL)和微晶颗石(Nannoliths, NAN)(自己瞎翻译的),异晶颗石是由复杂的晶体单元呈径向排列组成,同晶颗石是由许多细小的菱面体晶体通过有机基体组合成的,不符合以上两种形态的称为微晶颗石,现存的颗石藻类中,已经区分出180种异晶颗石,90种同晶颗石和10种微晶颗石。当然,从形态上分,还有其他一些难以归类的颗石类型。
颗石的形态和分类,用文字真是不可描述,还是直接上图好解释。
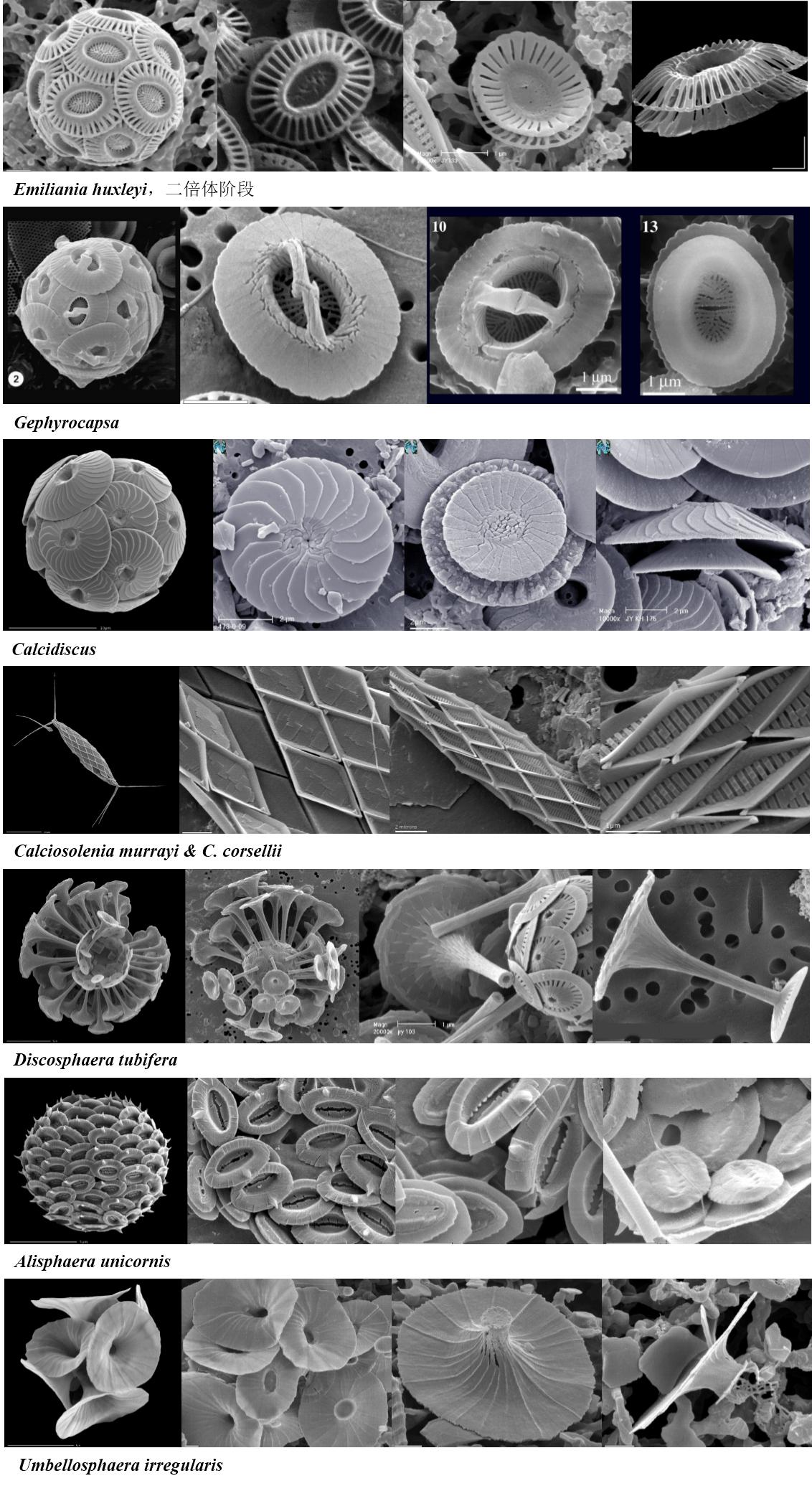
图4. 这组图为异晶颗石(藻),多数是盘碟状

图5. 以属Syracosphaera的几个种为例子,同晶颗石的颗粒特别明显

图6. 微晶颗石的形态往往是板状片状。
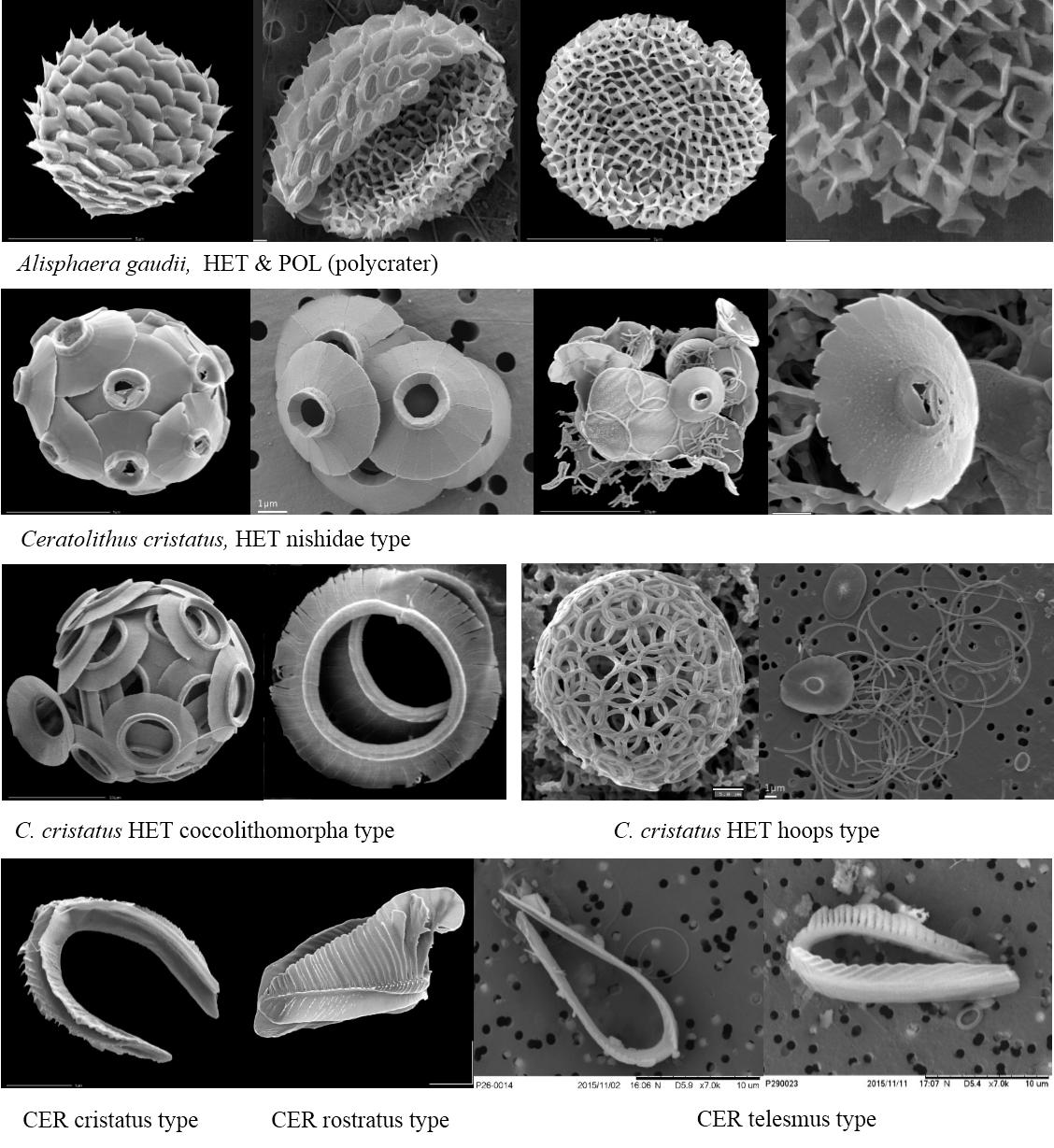
图7. 还有些颗石(球)不容易归类,如Alisphaera gaudii, 有异晶颗石的表型,还有种表型似密集的火山口状(polycrater,POL,文石而非方解石)。又如Ceratolithus cristatus,异晶颗石就有三种类型(西田?nishidae, 颗石翼状coccolithomorpha和箍形hoops),而该属还有一种马鞍形颗石(Ceratoliths, CER, 属微晶颗石?)
4. 生活史
定鞭藻类有单倍体-双倍体生活史,意味着两个阶段都有独立的有性分裂过程,并且有其独特的模式形态。如下图8. 颗石藻Coccolithus pelagicus,在单倍体及二倍体阶段均有二分裂法无性生殖,且两者的颗石形态不同,二倍体阶段为异晶颗石,单倍体阶段为同晶颗石,单倍体参与有性生殖(配子生殖)后,细胞则同时出现两种颗石。更多的研究发现,有相当种类的颗石藻存在此现象(如图9.),而且还伴随着细胞形状的巨大变化。
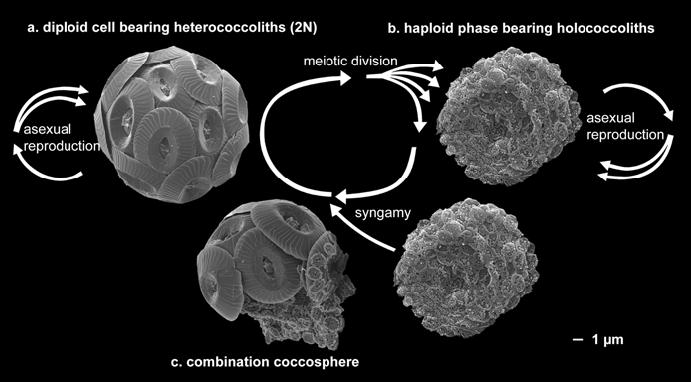
图8. 颗石藻Coccolithus pelagicus的生活史((Billard and Inouye 2004)
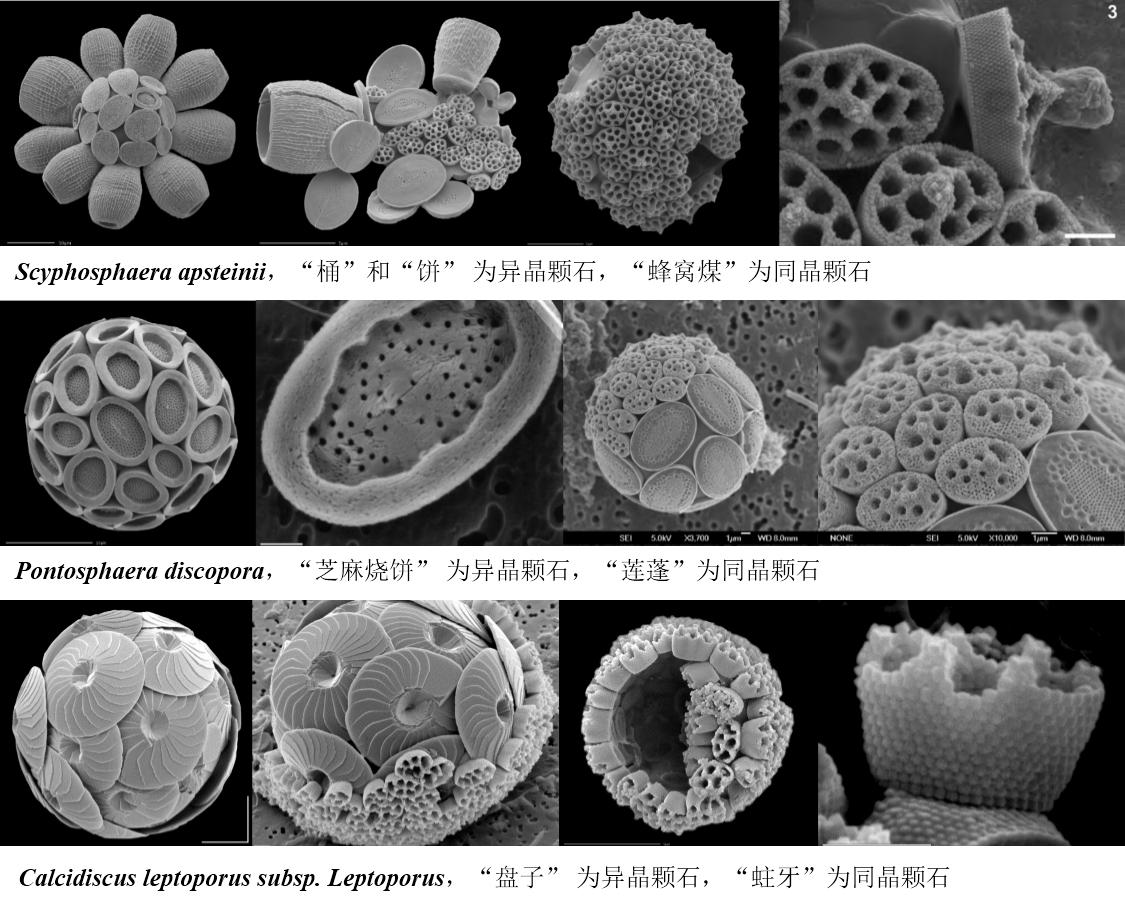
图9. 异晶和同晶颗石经常在同一种颗石藻中出现,通常异晶颗石是二倍体阶段的表型,而同晶颗石是单倍体阶段的表型,而且两种颗石可能出现在同一个细胞中(单倍体阶段)。
5. 颗石的形成
研究已经证明,异晶颗石是由细胞内的高尔基体分泌,然后转运到细胞体表面,但同晶颗石的形成还未能确定,有认为也是高尔基体形成,但尚未在细胞内观察到,但其他研究认为是在胞体鞭毛附近的表面或邻近位置发生的。等鞭金藻目(Isochrysidales)的种类形成钙质同晶颗石发生在内质网中的一个特殊液泡系统中,直接联接到核膜上,在单倍体阶段,不会形成钙质鳞片。环境中采集的Alisphaera(海石藻?)中发现了组合颗石型,具有文石(态)正方形微晶颗石,但这个特征的颗石藻还未有实验室培养的研究。
6. 异形世界
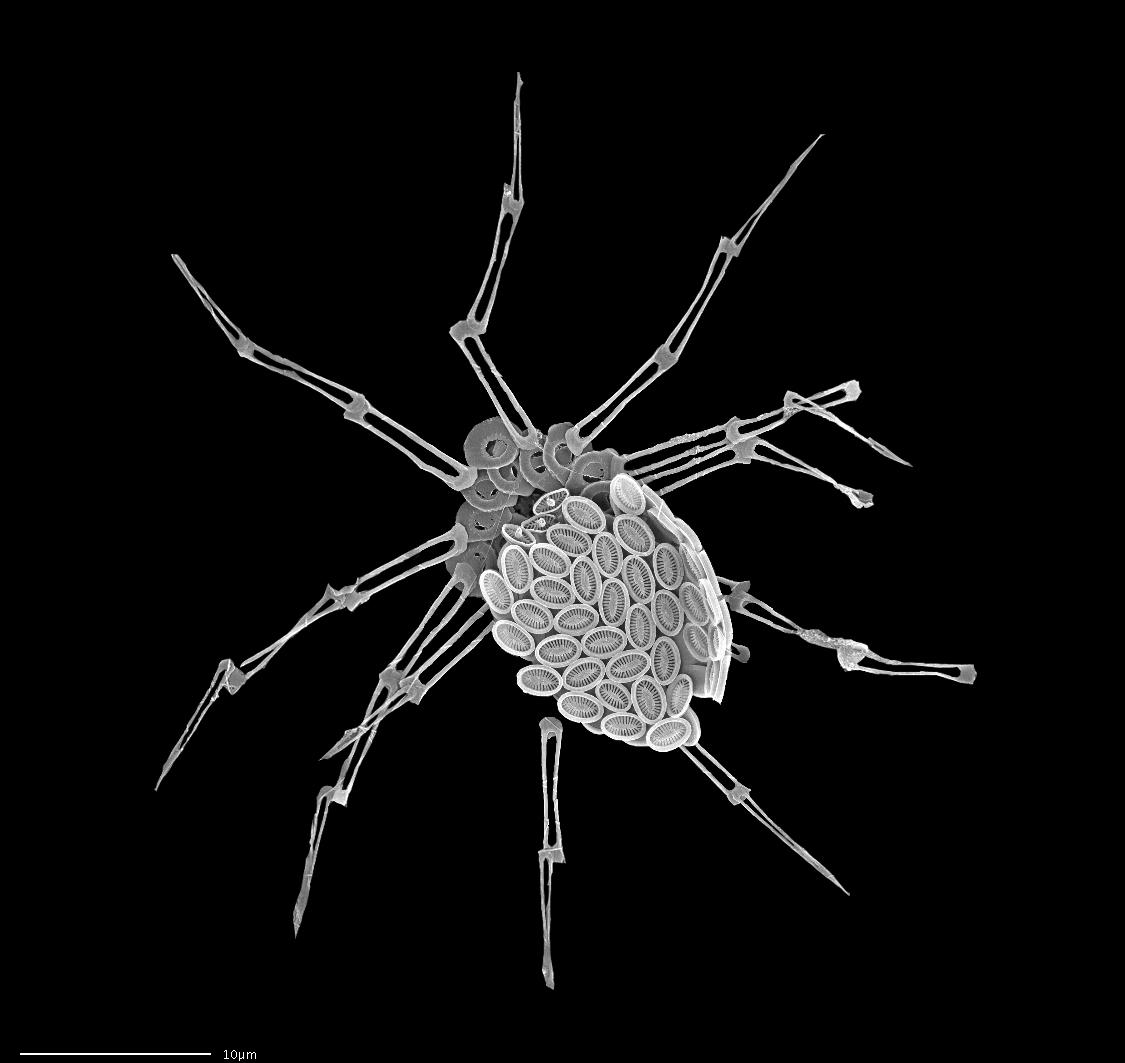
图10. 颗石藻Michaelsarsia elegans
这个图(图10.)绝对有星际大片的感觉,微藻一下子从大海走向了星辰。颗石藻M. elegans出现了变异形的颗石,称为附肢状颗石,就像蜘蛛的腿一样。
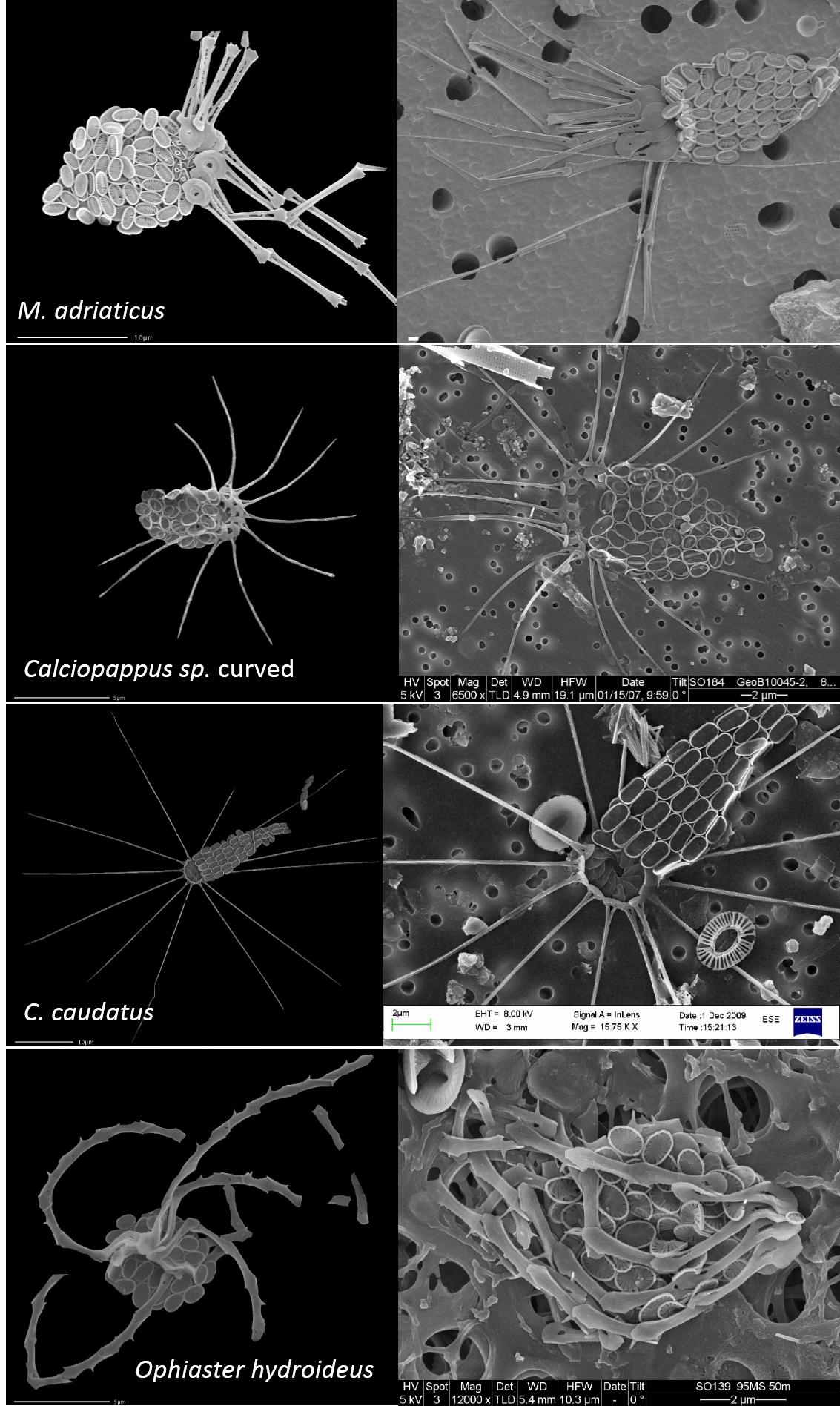
图11. 几种有附肢的颗石藻,看着真像动物……
M. elegans及图11中几种长腿的颗石藻均属于Syracosphaeraceae(条结球藻科?),另外如该科的属Syracosphaera,也有类似的颗石连成长链的情况。长长的附肢颗石,不仅可以张开,还能够收拢,甚至盘旋起来,如图12所示,再看看右下角的M. adriaticus,要说它是某种原生动物也没差啊?!
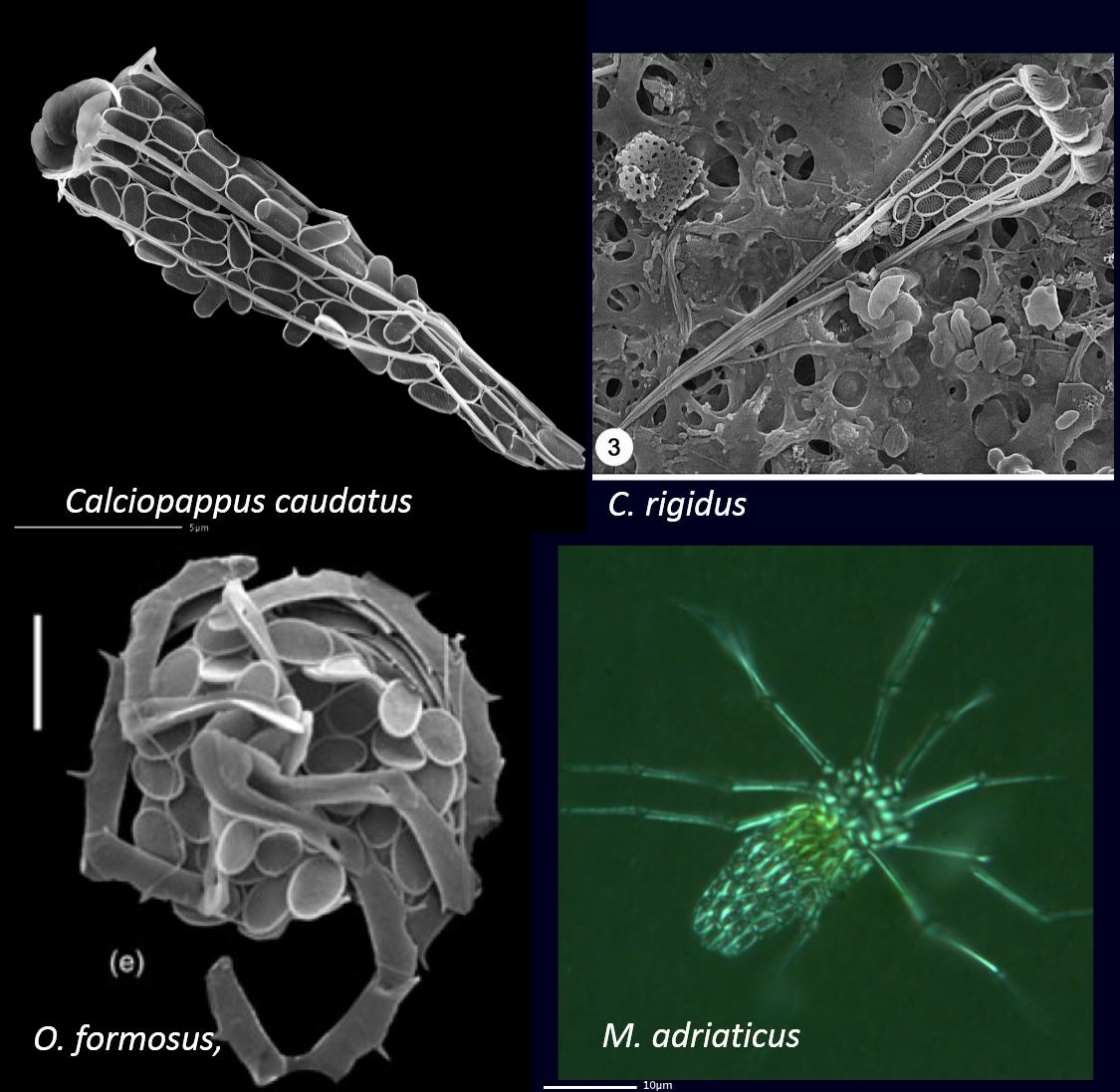
图12. 附肢颗石能放能收能盘旋。
7. 颗石藻的历史
因为方解石的稳定性,在海洋中又能够以较快速度沉降到海底,因此有机会很好的保留下化石来,可以说,颗石藻化石为研究地球尤其是海洋的历史带来了巨大的便利,也为我们了解颗石藻提供了详细的证据。
最早的记录,同晶颗石和异晶颗石分别出现在上三叠纪(~2.25亿年前)和下侏罗纪(~1.18亿年前),在三叠纪和侏罗纪交界期(约2.05亿年前)的物种大灭绝之后,贯穿整个侏罗纪和白垩纪,尽管有物种灭绝现象,钙质微化石的种类还是出现了一个增长,在坎帕阶和麦斯里希特阶出现了峰值(8000至6800万年前),然后突然消失了90%的种类(白垩纪-早第三纪大灭绝,陨星撞击)。有研究认为是陨星撞击地球,引起了海洋(硫)酸化,从而导致了很多海洋生物的灭绝,相对应的,现代发生的海洋酸化也已经造成了一些海洋钙质浮游生物的死亡,包括颗石藻。
少数存活的颗石藻种类在大消亡时期之后开始迅速的繁生。然而新的新生代种类又迅速出现,替代了前者(钙质微化石的多样性丰度在古新世晚期到始新世早期有所上升,而在始新世中期到渐新世又迅速降低,在中新世恢复又在上新世和更新世消减(如图13.)。
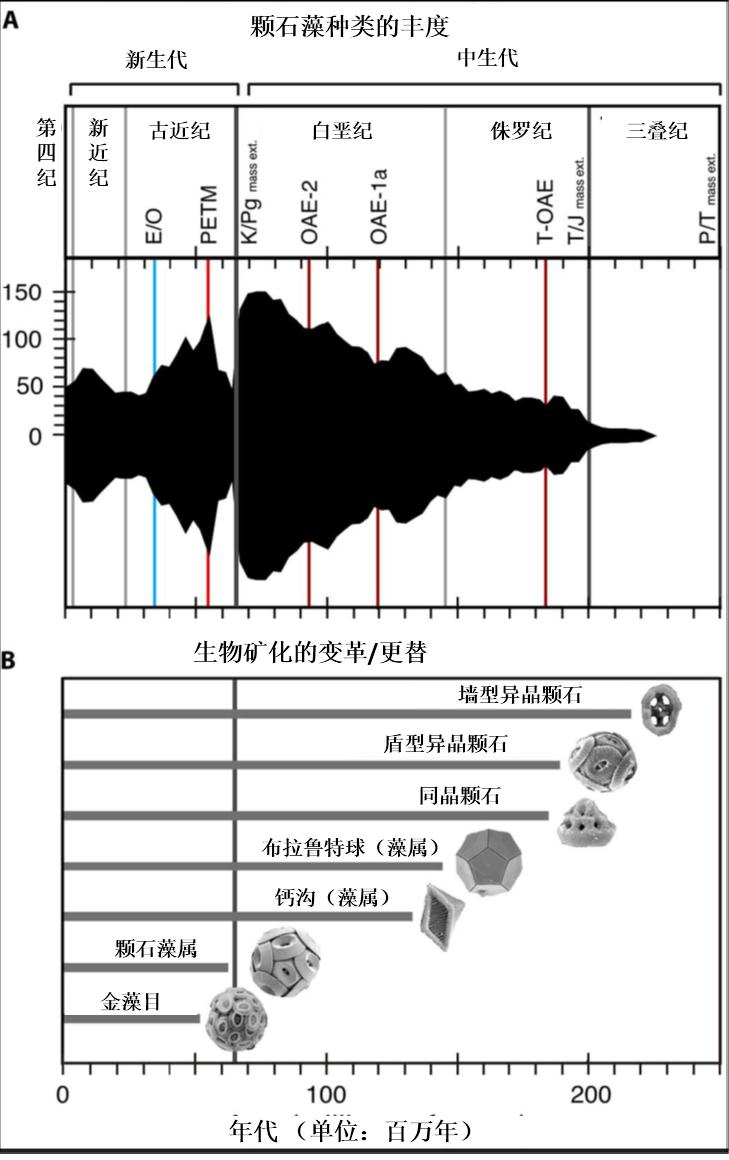
图13. 颗石藻的进化史(Monteiro et al., 2017)。(A) 颗石藻的种类的丰度。E/O,始新世/渐新世,冰川时代开始;PETM,世极暖时期(古新世/始新世);K/Pg,白垩纪/古近纪;OAE,海洋缺氧事件;T-OAE,托尔阶海洋缺氧事件;T/J,三叠纪/侏罗纪;P/T,二叠纪/三叠纪;mass ext,生物大灭绝。 (B) 主要颗石藻生物矿化更替和形态类群的化石记录,包括了墙型(异晶)颗石的出现(颗石具有狭窄墙型的环),盾型(异晶)颗石(盾板互相卡位构成结实的颗石球),同晶颗石(单倍体阶段出现,微晶体构成),布拉鲁特球(Braarudosphaera,布拉鲁特球藻属,五边形的片状微晶颗石层叠构成十二面体颗石球),钙沟(Calciosolenia,钙沟藻属,独特的菱形墙状颗石); 颗石藻属(Coccolithus,丰富而广泛分布新生代属种),金藻目(Isochrysidales,主要的颗石藻目属,包括了Emiliania, Gephyrocapsa和Reticulofenestra等种类),横线表示重要的生物大消亡事件及古海洋/古气候事件。
.
9. 颗石藻与古海洋环境重建
作为海洋沉积物年代标志物质,颗石藻的钙质微化石具有非常重要的意义,提供了可靠的年代标记,而且样品需求量少;不仅应用于学术研究,还为石油地质学所参考。钙质微化石的分类学及地层学演变等内容已经在过去的50多年中被广泛研究。一个有趣的现象是,同一种(群)内,微化石增大的趋势经常可见。
钙质微化石同样被应用于古海洋环境重建的研究。水平分布的水团变化与全球气候变化是相对应的,如“冰期-间冰期”周期变化。海洋表层水的不同水团中的颗石藻种类组成是不同的,同时海洋表层沉积物中的钙质微化石群体的组成反映了活性群体的组成。因此钙质微化石的地层学变化可以理解为等同上覆水团的变化,基于此理论,地质年代不同区域海洋表层水的结构组成得以分析重建。如冷水和温水水团中E. huxleyi 形态类型的构成是不同的,颗石直径>4.5μm的E. huxleyi在冷水团区域中较为少见,因此直径>4.5μm的颗石化石就可以用来界定地质年代冷热水团的区域。如金藻目(Isochrysidales)成员所产生的不饱和烯酮含量,可反映该物种生存区域的水温,烯酮具有对抗成岩作用的功能,可以从上溯到1.05亿年的海洋沉积物中获得,因此海洋核心沉积物中的烯酮含量可以用于分析古海洋表面的温度。又如光密度是低透光区域藻类(如F. profunda)生存最重要的限制因子,低透光区域光密度取决于光在(上层)高透光层的的透射程度,也就是取决于高透光层区域藻类的丰度,进而取决于该层的营养盐浓度及水层强度,因此依据低透光区颗石藻种类的钙质微化石相对丰度,可以分析地质时代表层海水的信息。

图14. 一些颗石化石,地质时期的各类颗石花样比现存的种类还要有意思(www.ucl.ac.uk)。
10.颗石的诞生与历史变化
(本节内容是搬运来的)
最早的颗石化石可追溯至2.25亿年,意味着定鞭藻类群的碳酸钙(方解石)生物矿化始于三叠纪晚期。定鞭藻的起源更早,分子遗传进化研究认为它们从其他藻类中分化出来的时间是新远古纪(12亿年前)。海洋浮游生物类群在这之间漫长的时期中没有出现矿化颗石,但是颗石藻及其碳酸钙生物矿化功能的出现,标志着一种近单向的多样化趋势的开始,同时也是地球历史上第一次大规模的碳酸盐沉积。随着时间的推移,对颗石岩的多样性的估计反映了在颗石藻结构上的形态创新和多样性的迅速积累,并显示了物种丰富的多样性,这是早期进化历史的特征。
这一趋势仅被一种独特而明显的瞬时环境扰动所打断,这种干扰与白垩纪-古近纪界线大灭绝事件有关(0.66亿年前),这个时期消失了90%以上的颗石藻物种,而后伴随着始新世和渐新世的冰川气候变化,又经历了长期的物种多样性下降。在过去的3千万年中,颗石藻类进化的总体趋势是随着物种的逐渐减少导致大量的高钙化颗石的逐渐减少。
尽管与较早的古近纪和白垩纪相比,这一趋势导致了今天的主要物种的颗石大小的减少,但现代颗石藻群体仍然保留了大量的颗石结构和细胞形状。从形态学上来说,所有的颗石藻都有相同的基本结构:外骨骼球团包围着细胞,但是细胞的形状有显著的变化,以及颗石的形状、结构、晶体结构、数量,多样性,以及细胞周围的排列方式。颗石球的形状范围由球形到圆柱形,尺寸从3到30μm不等。每个颗石球上,颗石的数量从6个到几百个不等,在单个或多个层面上都有不同,而颗石的形状从简单的圆盘状结构到有装饰或突出的突出物,包括长刺、喇叭状和烤架形状。此外,许多种类只会产生一种单一的颗石,而颗石球是由几种不同类型的颗石构成的。最后,在多倍体生物矿化模式中有一些变化取决于它们所产生的“单倍-双倍体”生命周期的阶段。

图15. 现存活于世的部分颗石藻种类(Monteiro et al., 2017)。 (A) Coccolithus pelagicus, (B) Calcidiscus leptoporus, (C) Braarudosphaera bigelowii, (D) Gephyrocapsa oceanica, (E) E. huxleyi, (F) Discosphaera tubifera, (G) Rhabdosphaera clavigera, (H) Calciosolenia murrayi, (I) Umbellosphaera irregularis, (J) Gladiolithus flabellatus, (K and L) Florisphaera profunda, (M) Syracosphaera pulchra, (N) Helicosphaera carteri。(标尺=5μm).
在二倍体阶段,颗石藻会产生异晶颗石,由大的、完全共生的方解石晶体的径向阵列形成。与此相反,在单倍体阶段,许多物种产生的同晶颗石,它们是由微小的(0.1μm),等轴分布的方解石菱面体通过一个有机基体构成。异晶和同晶颗石生物矿化起源于颗石藻多样化的早中生代早期。而不同的细胞形状,各种各样的颗石类型,以及(搭建)结构的多样性都属于颗石藻生物学的保守特征,可以通过颗石藻上百万年的生物矿化历史来识别的。这种变化的组合效应在颗石的形状和大小上,它们在颗石球上的排列,以及颗石球的形状和大小,在群体中产生了显著的形态多样性。这种形式的多样性和在颗石藻中形态特征的长期保守性,引发了什么是生物矿化的潜在优势这一问题。
11. 颗石藻之生物矿化
(本节内容还是搬运来的)
从最广泛的角度来看,矿化颗石(钙板)的产生很可能是保护细胞的需要,这是对多个浮游生物群体的挑战,例如硅藻形成硅质骨架,又如甲藻使用碳酸钙板和坚韧的细胞内有机板。然而,除了对保护罩的一般需求外,还可能会有一个由颗石球形态引起的更复杂的功能,这是由环境类型、颗石球特征和颗石(搭建)结构之间的广泛的生物地理联系所证明的。

图16. 不同环境下的颗石藻种类,其细胞和颗石有明显的区别。
例如图16.所示,寡营养海区的两类典型颗石藻,U. irregularis 和D. tubifera都是不动型颗石藻,拥有大而低密度的球体结构,特征是形体大、喇叭状的颗石围绕着小小的有机团。中等营养和富营养环境下,占主导地位的是Gephyrocapsa (包括E. huxleyi),C. pelagicus, 和 C. leptoporus之类带有扁平碟状(扁盘状)颗石覆盖的颗石藻种类。较深的亚透光层海水环境中占主导地位的是F. profunda和G. flabellatus等游动型颗石藻,球体相对较小,颗石为非常有特点的刀片状。
特定的生物地理环境与不同的颗石球结构相对应,暗示了颗石的形成与生态适应之间的潜在联系。E. huxleyi对分布区域的环境控制以及响应环境变化等生态学内容已经有了很好的了解,但其令人好奇的复杂性和多变的颗石类型仍然引发了关于为什么颗石藻会出现生物矿化的争论。
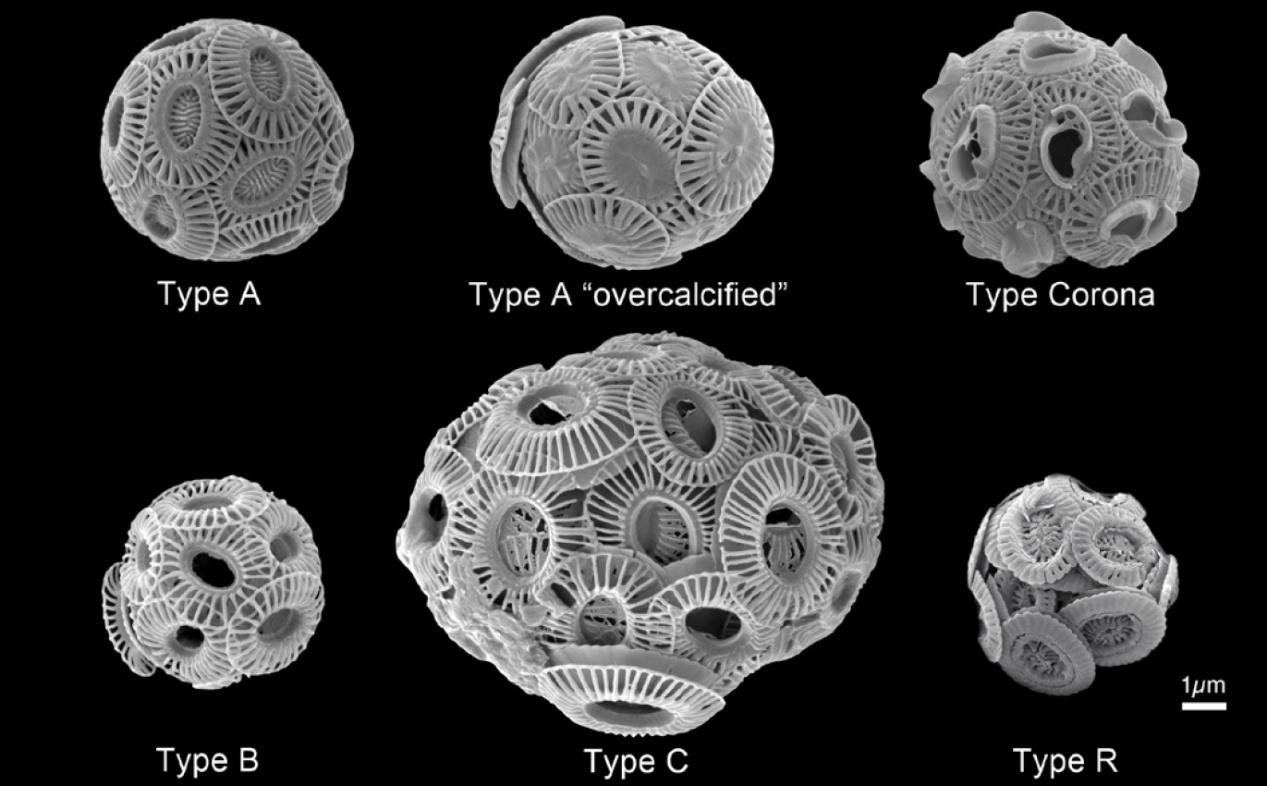
图17. 赫氏颗石藻(E. huxleyi)在不同的环境中,细胞及颗石具有多种表型(引自phys.org)。作为一种广布种,随分布海区的不同不仅其颗石表型有差异,而且另有研究发现,其基因的多样性也是相当可观。
12. 生物钙化的作用
对于颗石藻生物钙化功能的产生或者在遗传上保留这一特征的原因,有过很多的分析推导,某些观点认为钙化这一功能可以给藻体带来某些益处。
(1)促进光合作用。有观点认为钙化是为光合作用的碳浓缩机制服务,它能够降低细胞周围的总碱度,提升CO2分压。而研究还存在很多争议,从所有的实验证据看,钙化并不直接形成碳浓缩服务光合作用,大多数颗石藻种类在颗石完成单层或交叠覆盖细胞时,即停止了钙化,而且钙化与光合作用的碳供应相竞争。另一个说法是,颗石的存在具有散射光线的作用,提高光照射到色素体的几率,从而提高光合作用的利用率。有许多研究证据支持这一点,尤其是在光缺乏的海洋的深透光层,这一能力对颗石藻意义很大。如F. profunda(颗石结构似雷达盘)和 G. flabellatus(50-150米深度)(见图16.)。
(2)光伤害保护。对于生活在上层海水的颗石藻而言,钙化具有保护细胞免受光伤害的功能。强光条件时,颗石钙板的存在可以为细胞挡光(尤其是紫外光)或者散发掉能量,而且细胞表面颗石层的数量还会根据紫外的强弱进行调节。
(3)流体动力控制。浮游生物生活在海洋表面,通常是寡营养环境,而在深处通常是富营养,颗石的密度较高,可以起到“压舱石”的效果,将藻细胞沉降到深处,便于吸收营养,沉降的速率可达十倍的效果,只不过,相比其运动型种类的运动速度,这个效果并不算得上可观。
(4)抗侵染。这一点非常具有争议性,这观点认为,颗石球的“盔甲”结构可以保护藻细胞不被侵入细胞而导致死亡,例如某些病毒会感染E. huxleyi,甚至能终结这类藻华,病毒的入侵需要穿过颗石球壳达到细胞膜,钙化机制的存在可以一定程度上抵抗病毒感染。
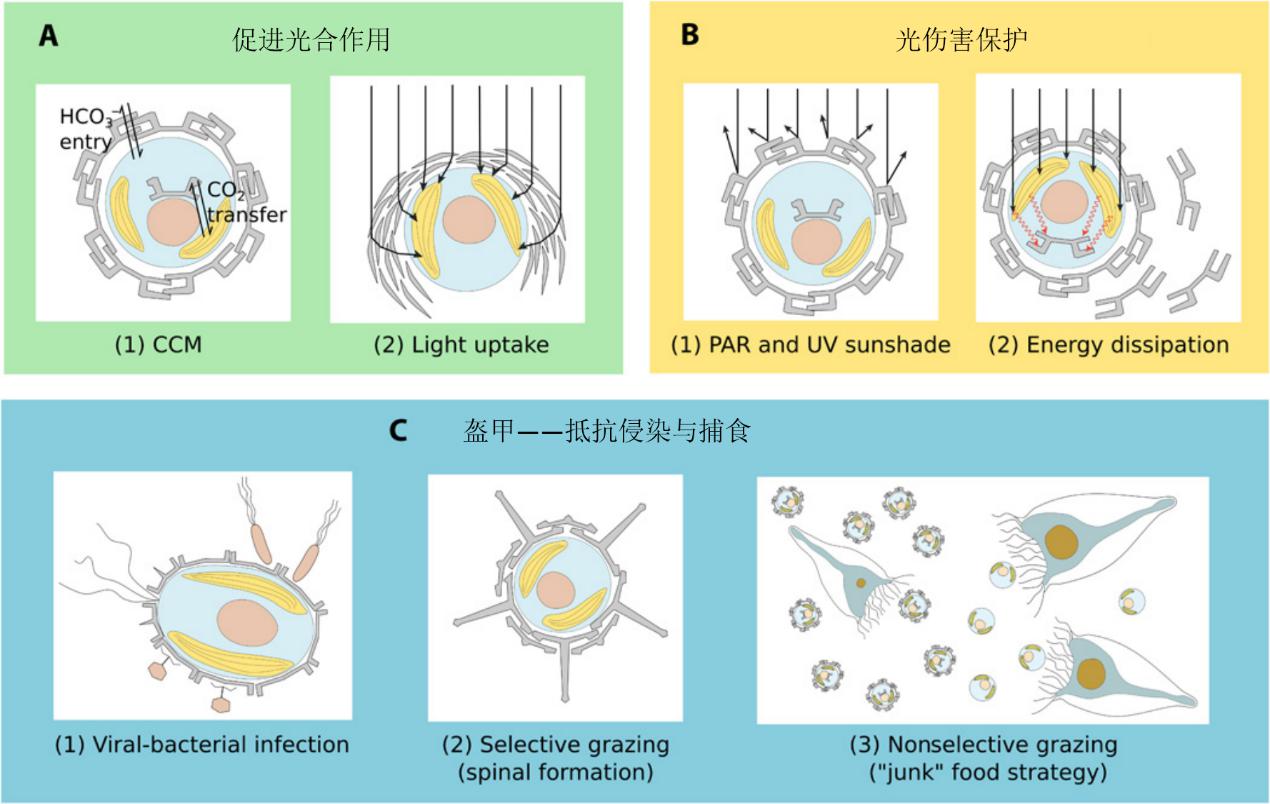
图18. 生物矿化可能的有益作用(Monteiro et al., 2017)
13. 颗石藻与生物地球化学循环
从颗石藻这个类群诞生(约2.25亿年前)到进化至今,大约有4000多种形态多样化的种类灭绝了,现存发现的种类约280种,90种是同晶颗石藻,其余的190种中(不包括90种同晶颗石藻),仅有2种能够在大洋中形成藻华,即赫氏颗石藻E. huxleyi 和大洋桥石藻G. oceanica。

图19. 卫星拍摄的1999年英国康沃尔(Cornwall)南部海域爆发的赫氏颗石藻(E. huxleyi)藻华(白水现象)(提供者Steve Groom)。
颗石藻广泛分布于各种环境下的海洋表面,从高生产率的温带富营养区域和近极地区域到长久贫营养的亚热带区域。在近极地、温带和热带,颗石藻大约贡献了1%到10%的基础生产力/生物量,在形成藻华的情况下,可增加到40%。与有孔虫一样,颗石藻是地球上最旺盛的浮游钙化生物。
它们持续的产生碳酸钙“雨”,降落到深海,保持海水垂直梯度碱性(碳酸盐泵角色),“颗石雨”建立了地球最大的碳元素地质表层,而海床碳累积物对碳循环的敏感性在地球气候稳定中产生重要的稳定反馈。而且致密的矿物颗石在将有机物传输至深海中扮演了压舱石的角色,因此也对海洋CO2垂直梯度做出了贡献。颗石藻的重要贡献在于调节海洋生物地球化学循环和气候,需要我们充分了解它们的生理学和生态学功能,并将之应用于调节海洋生物地球化学循环。
海洋是地球诸元素生物地球化学循环的关键组成部分,而且代表了大气和岩石圈之间一个重要的交互界面。作为一个主要的浮游生物群体,颗石藻通过吸收并加入有机和无机物质或通过气体释放,参与到几个元素的循环中(如氧气和二甲硫醚)。生物活性元素(如碳、氧、氮、磷、硫和铁)在短期(如几天到千年)和地质时间(百万年)尺度上循环。高度海洋滞留时间的元素,如钙和镁(分别为1百万年和1.3千万年),主要由于海底传播速率的变化而改变了浓度,而非地质时间尺度。
对颗石藻而言,碳和钙的循环最为突出,两个循环在地质时间尺度上都是紧密联系在一起的。
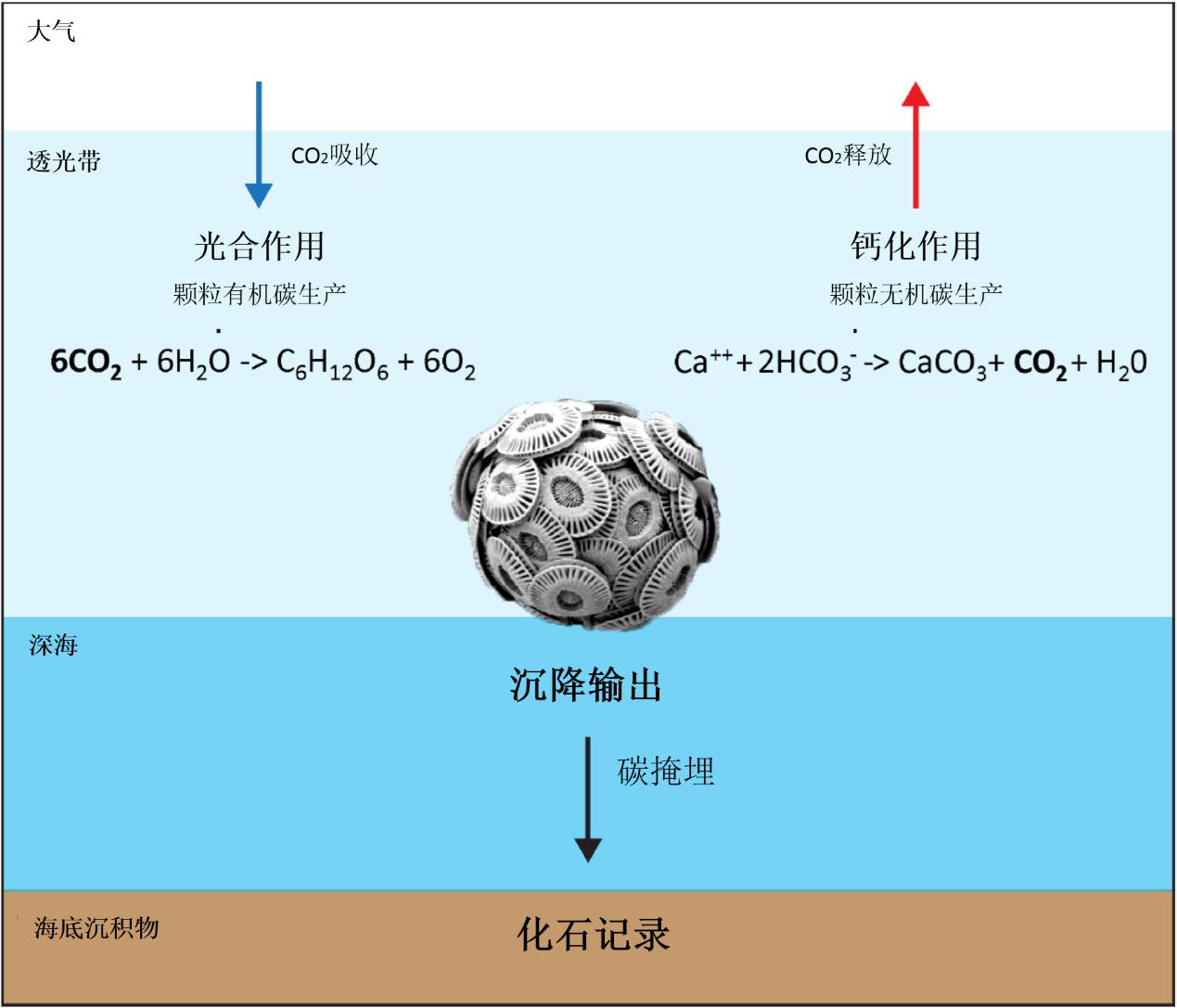
图20. 颗石藻在生物地球化学碳循环中的示意图(Luka Šupraha)
(一)碳循环
自颗石藻诞生的2亿多年来,它在地球系统的无机和有机碳的命运中扮演了多重角色。全球的碳元素储存在不同的储层中,且它们在不同的时间尺度上相互配合。地球最大的碳储层是的岩石层,含6亿 PgC(1015克碳);其次是海洋,拥有4万 PgC(主要是可溶性无机碳,DIC);两个储层由含有150PgC的沉积物连接起来。整个地球生物圈占有2000 PgC,大气圈再增加750 PgC(主要是CO2形式)。当前大气中CO2浓度大约是380多ppm,且因为人类活动如化石燃料燃烧、森林砍伐、水泥生产和土地开发等原因,每年增加约3.3 PgC。在短时间尺度上,岩石圈对系统中碳的通量可以忽略不计,因此海洋在碳的命运中扮演着主导角色(取代岩石圈,成为最大的碳储层)。
颗石藻是在上层海洋的光合作用区域内对CO2的光合作用起到贡献,以及将有机物质排入到深海中,这就导致了海洋中CO2的吸收,被称为有机碳泵(输入海洋)。另一方面,碳酸钙的生物生产和排放抵消了CO2的光合作用通过改变海洋碳酸盐系统来实现CO2的净释放,成为无机碳反向泵(输出到大气)。输出的无机碳和输入的有机碳降雨的比率(Cinorg/Corg)称为“Rain ratio”(降雨率?)。生物界的部分物质会被重新“唤醒”,在海洋中浮游植物也会再次出现。从上层到深海的CO2的强度是由“Rain ratio”所影响的,因此,通过将碳排放到深海,“Rain ratio”改变了大气与海洋表面的CO2浓度。颗石藻在“Rain ratio”中扮演着重要的角色,不仅是因为生产有机颗粒和无机碳,还因为其高密度的颗石(方解石)产生的压舱石效应。方解石丰富的颗粒在沉积速度和沉积速度提升的同时,还可以在深海中沉积。因此,钙质的“海雪”可能会增加有机碳泵的强度。

按照科学家的研究计算,颗石藻引起的碳酸钙的沉积作用(沉积速率大于溶解速率),每千年可累计10mm-100mm厚度。海洋沉积物形成了海洋和岩石圈之间的界面,约48%面积的海床上在持续发生含碳酸钙物质的沉积或埋藏行为。
板块构造运动在俯冲带的位置将海底置于大陆边缘,在地球深处,古海洋沉积物形成碳酸盐岩石,主要是石灰岩、白云岩[CaCO3和CaMg(CO3)2],以及化石燃料。CO2从岩石圈释放出来主要由火山活动和部分非火山排气释放,另一个作用是地壳构造的力量把以前埋藏的碳酸盐岩石隆起到地表,在那里它们暴露在风化层中,碳酸盐岩的化学风化从大气中吸收CO2,并通过河流向海洋释放成分离子(如Ca2+、HCO3−、Si4+、Mg2+),这些离子被浮游植物所利用。人类直接通过化石燃料产生的CO2直接影响自然碳循环,而化石燃料是由地质时代产生的。

(二)钙循环
钙是现世海水中第五丰富的溶解离子,并被认为“保守”,这意味着钙离子在海水中的浓度不会受到温度、pH值和压力的变化而影响。据信,海洋的钙循环在地质时期处于稳定状态,但在过去几十年里发表的论文揭示了事实并非如此。钙的生物地球化学循环与无机碳循环紧密相连并与控制海洋碱度和大气CO2的过程相结合。
海洋中钙的浓度是由几个过程的平衡控制的,下表1中进行了总结:(1)生物源性介导的碳酸钙的形成和埋藏,从海洋中除去钙;(2)热液过程(hydrothermal process)和(3)白云石化(dolomitization)是Ca2+的来源,以及(4+5)含钙岩石的风化和随后的河流输入。其中颗石藻生物钙化功能在此起了重大的作用。
表1. 钙元素的生物地球化学过程
| 生物源钙化 | Ca2+ + 2HCO3- à CaCO3 + H2O + CO2 |
| 热液过程 | Mg2+ + Ca−basalt à Ca2+ + Mg−basalt |
| 白云石化 | 2CaCO3 + Mg2+ à MgCa(CO3)2 + Ca2+ |
| 碳酸盐风化 | CaCO3 + H2O + CO2 à Ca2+ + 2HCO− |
| 硅酸盐风化 | CaSiO3 + H2O + 2CO2 à SiO2 + Ca2+ + 2HCO− |
14. “带壳”的海洋浮游生物
回看本文图3.,有个黑色三角形标记的藻叫Hyalolithus neolepis,这个藻并不在颗石藻的范围内,因为它的生物矿化不是钙矿化,而是跟我们所熟知的硅藻类似,存在硅矿化现象。
海洋中,除了颗石藻,有孔虫、甲藻和硅藻之外,还有些类别的藻类或者浮游动物也有类似的披甲带壳的现象,这些生物的矿化或带壳功能如何进化得来,相互间又有什么关系,尚属悬念。本文就放几个图片,让大家稍微了解下,谁让它们长得漂亮呢?
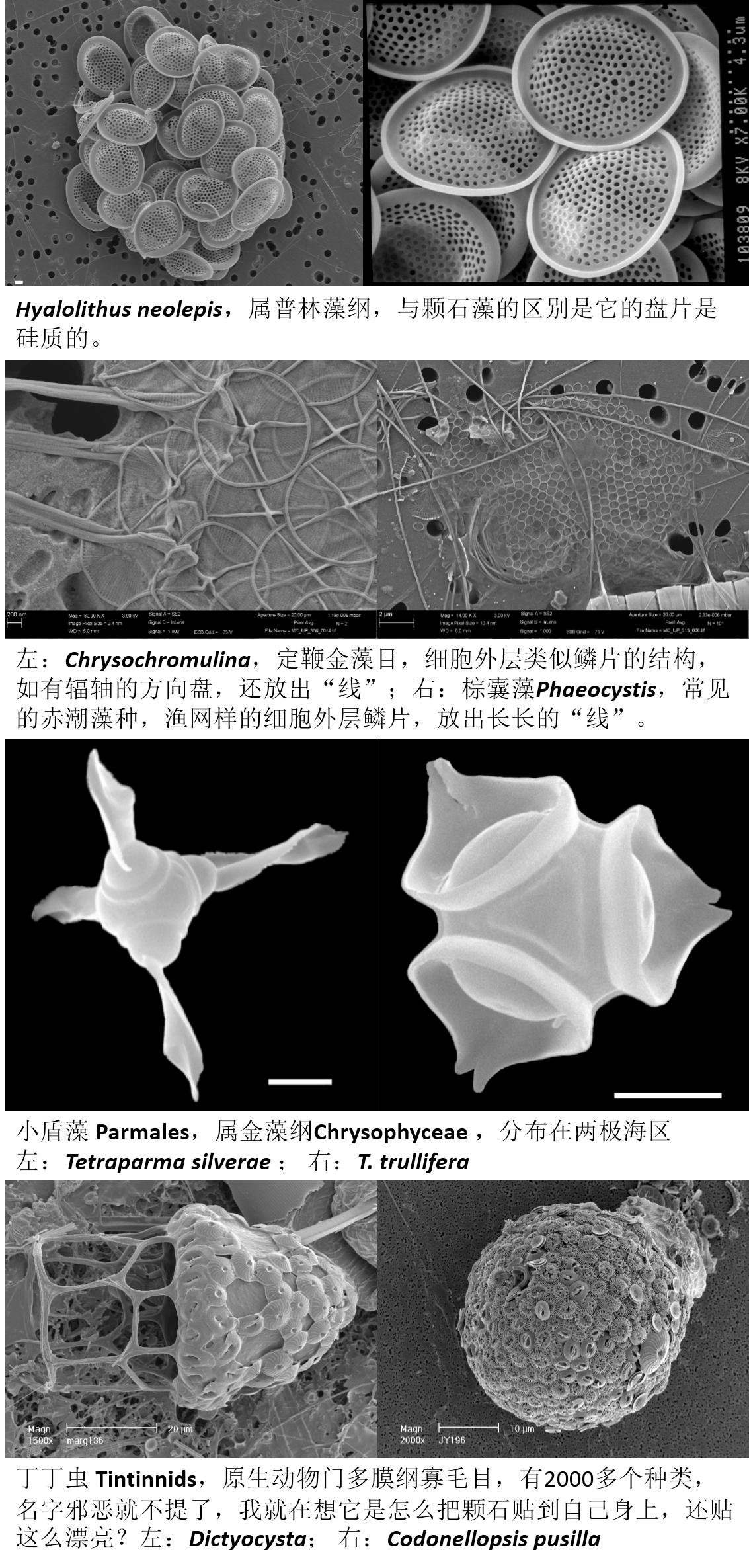
图23. 定鞭藻门下的几种非钙质的“带壳”种类以及披着颗石外皮的虫。
15. 颗石藻的未来
人类活动导致了温室气体浓度日益增加,在提升海洋温度的同时改变了海洋化学(组成),并有可能进一步对海洋生态系统造成广泛的影响,包括颗石藻的钙化度和生产率。
亚南极-南极区域的颗石藻地理生物学研究发现,在过去25年里,赫氏颗石藻E. huxleyi的分布区域向极点扩展,在1990年前赫氏颗石藻只分布在南极极面(南极圈范围)的北方区域,而现在开始向极点方向拓展。推测极地海域表层水温的上升和/或贫营养化导致了赫氏颗石藻向极地海域的拓展。
自工业革命以来,大气中增加的CO2约有30%被海洋吸收。导致了海洋表层水的pH值从8.18降低到8.06,预测到21世纪末,海水的pH值将进一步降低0.3至0.4。在未来海洋酸化将显著影响具有钙质骨骼的海洋生物(如珊瑚和有孔虫)的钙化,而对于颗石藻而言,因诸多培养或观察的研究结果未得到一致结论,持续的海洋酸化是否会影响颗石藻依然在争议中。有些研究发现高浓度的CO2会导致钙化程度降低,畸变程度增加;又有其他研究发现CO2浓度变化对颗石藻的钙化影响因种而异,甚至因株系而异(如E. huxleyi和G. oceanica种内遗传差异就非常大)。

图24. 不仅仅是E. huxleyi和G. oceanica的种内形态及遗传多样性高,Umbellosphaera tenuis的颗石也是模样多多(从上往下分别是Ⅰ、Ⅱ、Ⅲa、Ⅲb、Ⅳ和O型)。
颗石藻对海洋酸化敏感的差异认为是由遗传易感性导致的。实地观察发现,碳酸盐物质能强烈的影响颗石藻的地理分布,某些颗石数量通常与pH值呈正相关关系,而低pH区域的E. huxleyi具有高度的钙化颗石,某些地质时期化石样品的研究结果也与之相一致。因此,海洋的持续酸化,可能会导致颗石藻基因型、表型及种株的组成变化,以适应低pH环境。因此未来海洋酸化对颗石藻的影响尚无法确定,需要全面的了解现存颗石藻种类的生理学和遗传多样性信息,以及地质时期颗石藻对全球环境变化的系统反应。

图25. 上排是C. leptoporus,下排是C. pelagicus,左边第一个均为环境中采集的样品为对照,后面三个为用不同CO2浓度进行处理的实验样品,可以发现随着CO2浓度的增加,颗石发生了很大的变化,如果持续的海洋酸化加强,至少对某些颗石藻的影响可能是致命的(Langer, 2005)
化石燃料燃烧产生的CO2对海洋的影响是多重的,如提升了表层海水温度,营养迁移到深海的阻断效应等,对颗石藻影响也不是一言可定之,如温度增加对藻的影响,水体中可利用CO2浓度的上升,同时pH下降,温度导致海洋水层变化进而影响了光照强度。这些因素的综合,对颗石藻是利是弊,尚无定论。
(本文引用图片如非特殊说明,均来自http://www.mikrotax.org/)